
Pages 62-91
43. Canker on red oak associated with Strumella coryneoidea. These cankers often are abundant on young trees and seriously alter new stands of oak. The fungus may cause some decay of the wood. The fertile, sexual stage of the fungus, which occurs after the tree breaks off at the canker and falls to the ground, is Urula craterium (low right). The fungus infects the bark first, then the tree produces wedges of wood into the bark (arrows, upper right). The fungus later spreads around the wood wedge and the seesaw interaction continues until the tree is killed, or the tree breaks at the canker. But in some cases, the tree walls off the fungus so effectively that the canker is stopped.
Pages 62-63
44. Chestnut blight is caused by Endothia parasitica (Cryphonectria parasitica). After the fungus kills patches of bark, some still-living portions of the cambium may "explode" with new growth of wood and bark as shown here. Another type of response to the chestnut blight fungus is the production of pillars of wood that "reconstitute" beyond the infected and killed bark. In a sense, the infected tree begins to form a "new tree" beyond the infected tree. Trees have marvelous ways of survival, built upon recognizing the injurious agent and walling it off. Then the tree goes on to generate more healthy tissues beyond the infected and dead tissues.
Pages 64-65
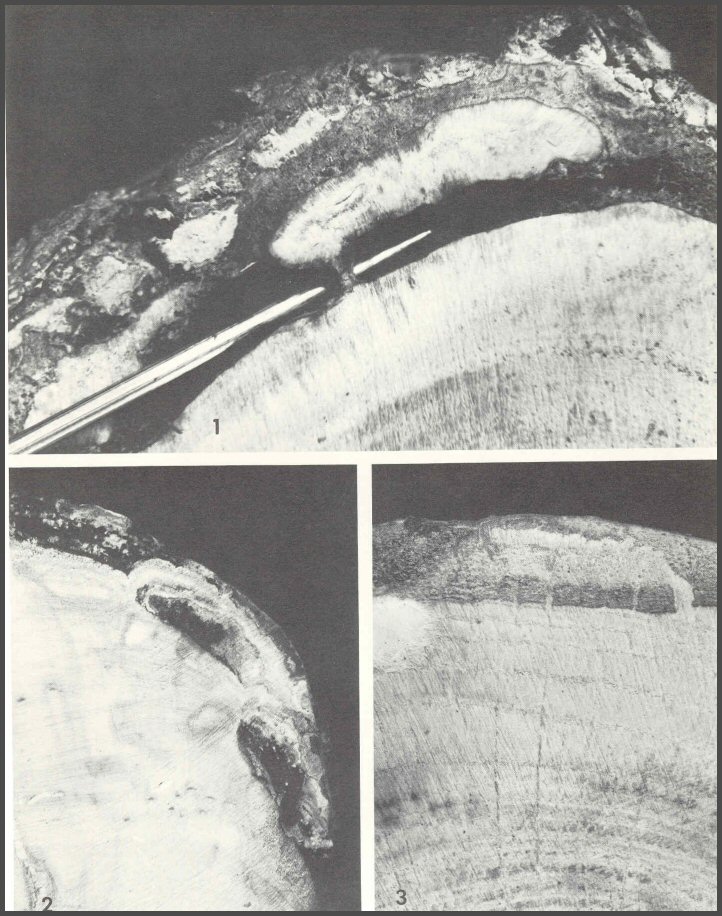
45. Wood wedges into bark: 1) American chestnut infected by Endothia parasitica, 2) Aspen infected by Hypoxylon mammatum, and 3) red oak infected by Strumella coryneoidea.
Pages 66-67
Page 68 = Blank
Interlude
To this point, many disjointed facts have been shown in the photos. No attempt has been made to connect them. The photos do give an historical account of the research. The keen observer will see in the photos much more than what is described. The keen observer will also start to perceive the sense of order that awaits and demands synthesis. If the microorganisms could move at will, they have had millions of years to do so, and trees as we now know them would not exist. If trees were able to stop all microorganisms, we would not find the many cankers and rot columns in trees that we do. Somehow, somewhere in time, some ecological evolutionary survival "deals" must have been made. We must recognize this. The basic rule: no absolutes! Trees do not "win" all the time. Yet, both must win enough of the time to assure the survival of the individual, and the survival of the group, long enough to adapt and to reproduce. What first appears as a paradox was resolved. The apparent paradox: Trees survive so long as they can avoid being digested by microorganisms. Microorganisms survive so long as they can digest trees. Both trees and microorganisms face a common survival threat - the environment. Again, an apparent paradox for environment was both very beneficial and very destructive - not only a threat, but a friend. It is the apparent, or accepted paradox that makes natural systems difficult to understand. Once we begin to understand the multidimensional mix of factors that both help and hurt, the order of things will become apparent.
The next group of photos shows this in the results of wounding experiments, and in further dissections that show the high order of the tree survival system, and the order of the survival system of wood - inhabiting microorganisms. The concept of compartmentalization and of microorganism succession began to emerge. The paradox is resolved: Microorganisms survive so long as they infect in the orderly sequence that best fits the survival of the group - succession. Trees survive after injury and infection so long as they recognize and wall off the infected tissues-compartmentalization.
Compartmentalization is a boundary-setting survival process. Boundaries are not absolute. When boundaries are weak, or when the force of the infecting microorganisms is very strong, the boundary will fall. To survive, microorganisms and the tree must win a time game with the environment.
Page 69
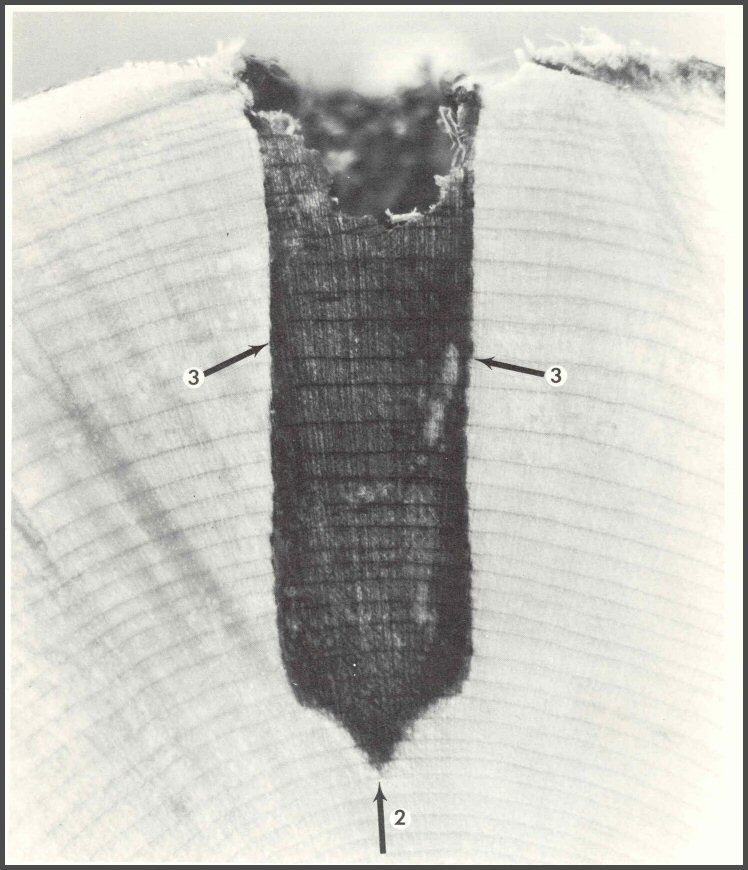
46. When a tree is wounded, as this red maple was with a deep drill bit wound, the tree responds by compartmentalizing the injured and infected wood. The wood is made up of many compartments. Each wood cell, with its strong cellulose wall bonded with lignin, is a compartment. Many small boundaried bodies or compartments exist within the living wood cells, especially the thin-walled protoplasm-containing cells, the parenchyma. The growth ring is also a compartment. The living strands of protoplasm that form a web of connections among the living wood cells are called, collectively, the symplast. When wood is injured and infected, the symplast connections between the infected and still-healthy wood are broken. Many chemical reactions take place to set firmer boundaries to resist, not stop, the spread of the infection. This is compartmentalization. It is a boundary-setting process to resist the spread of infection. It is not an absolute process. A model of compartmentalization has been developed. It is called CODIT, an acronym for Compartmentalization Of Decay In Trees. The model has two parts: Part I has three "walls" (two of them shown here). Wall 1 resists, not stops, vertical spread; Wall 2 resists inward spread, and Wall 3 resists lateral spread. Part II has one wall: Wall 4, which separates the infected cells from the healthy cambium. Wall 4 is a model of the barrier zone; it forms only where there is still living cambium.
Pages 70-71
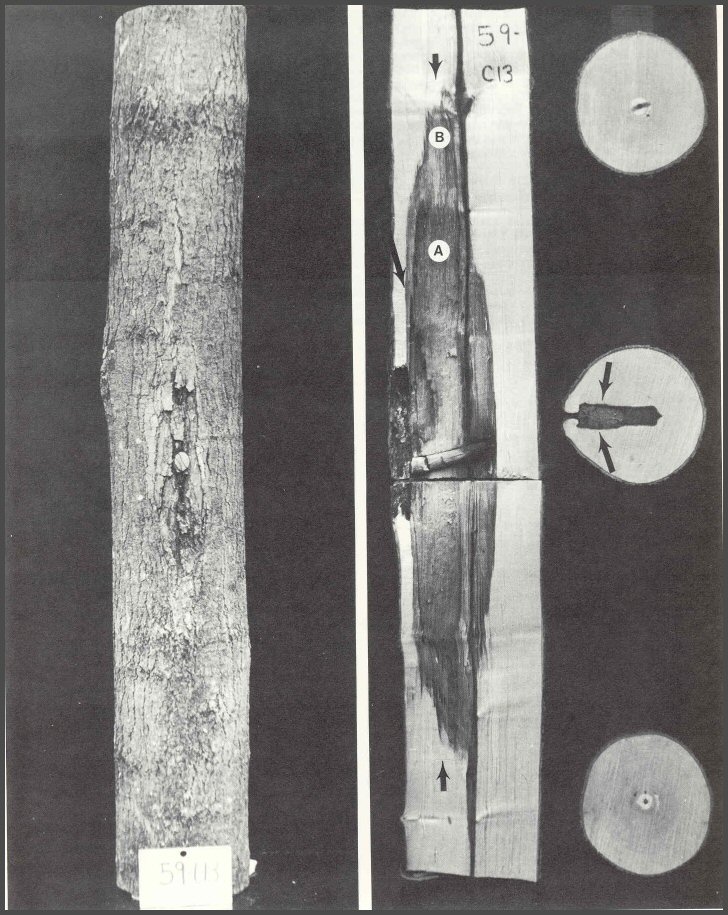
47. Sugar maple 59-C13 was wounded and a sterile wood dowel was placed in the drill hole. After 8 years it was dissected. The length of cambial dieback above and below the drill hole is shown by dark callus tissue. When trees are wounded, the cambium usually dies back above and below the wound and sometimes the dieback will extend to the sides of the wound. The three disks were cut from the ends of the bolt and from the center directly below the drill hole. Note that the end disks have only a very small central column of discolored wood, but there is a large column of discolored and decayed wood associated with the wound. The center disk shows that the shape of the column was approximately the shape of the drill bit. Note that decay started first in the younger wood tissues, contrary to what is stated in the textbooks. The arrows in the disk represent Walls 3 of CODIT. The large arrow shows the position of Wall 4, or the barrier zone. The small arrows at the ends of the column represent Walls 1 of CODIT. Wood in area A is decayed, while wood in area B is discolored. Wood chips from area B yielded bacteria and non-decay-causing microorganisms, while wood from area A yielded mostly decay-causing microorganisms. Because the wound went through the pith, Wall 2 was not operating. Wall 2 only resists spread inward to the pith.
Pages 72-73
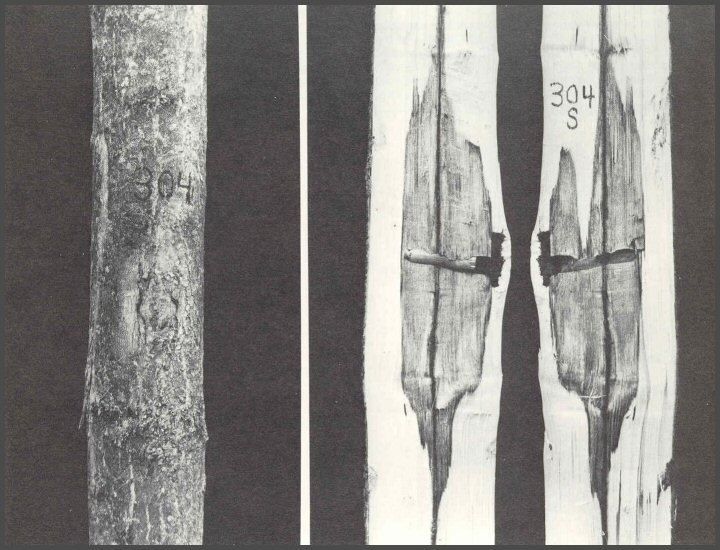
48. Tree 304S had no decayed wood associated with the wound. The column was very short. In the dowel experiments, many dowels were preinoculated with several decay-causing fungi. After 8 years, only a fourth of the introduced fungi were recovered. But, even when they were recovered, the decayed wood was within the outer column of discolored wood that yielded, on isolation, bacteria and non-decay- causing fungi. The concept of microorganism succession does not mean that bacteria ALWAYS came first, then non-decay-causing fungi, then decay-causing fungi. Definitely NOT SO! Succession means the orderly sequence of microorganisms. The order is such that the survival of individuals and groups is maintained in relationship to or in association with the specific environmental and other conditions. In some cases, it is possible that decay-causing fungi will be the first microorganisms to interact with the tree. Natural systems do not come in absolutes.
The wounding and inoculated dowel experiments were done in oaks also (also black cherry, white pine, and American beech). When heartwood was wounded it discolored. And the shape of the discolored column as viewed on the cross-sectional face of the central disk was the shape of the drill bit. The oak heartwood did compartmentalize the injured and infected wood. But, is not heartwood a dead, non- responsive tissue? It may be dead according to our definition, but it must be responsive, or at least reactive, to wall off the injured and infected wood.
Pages 74-75
49. When wounds or dying branches expose the wood of oaks to microorganisms, the sapwood begins to decay first, not the heartwood. This section of an oak shows the decay of the sapwood. Note also that the tree begins to wall off the decay at the base of the branch. When large oak branches die, and the central core is heartwood, the sapwood-rotting fungi decay the sapwood rapidly, thus leaving a core of strong heartwood. When such branches break, they do so a meter or more out from the trunk. When oak branches die before large cores of heartwood form, they usually break close to the trunk.
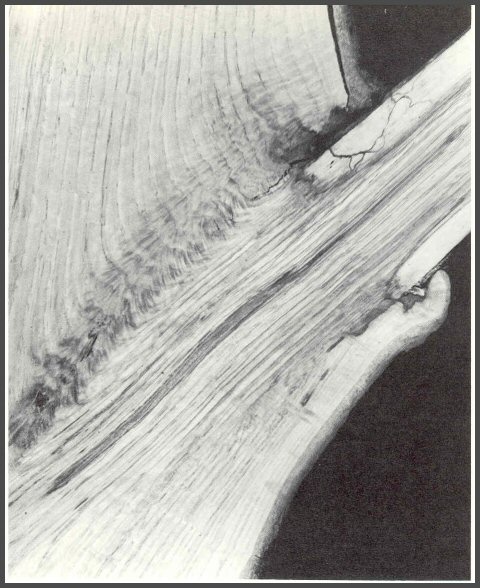
50. Other studies on red oak showed conclusively that heartwood does discolor, and that some of the changes that take place in the discoloration process are not so different from those that occur in maples. The infections did not spread into the center of the tree. The injured and infected wood was compartmentalized. But when other oaks were killed by basal girdling first, and then wounded, there were no sharp boundaries showing the outline of the drill bit in the heartwood. These studies indicate that heartwood will react when injured and infected. The exact nature of the reaction is still not clearly understood. We know only that it is orderly; it repeats. Also note that decay starts at the inner side of the sapwood and not deep in the heartwood.

Pages 76-77
51. Examination of naturally inflicted wounds and other wounds, such as this buckshot wound on a red oak, shows that the same principles are functioning, and the CODIT model is applicable in heartwood. Note the margin of discolored heartwood surrounding the decayed wood. Note also that the decay did not spread outward into the heartwood that formed to the outer side of the decayed wood. If the fungi are "heartwood rotting" then why don't they spread into the heartwood only one growth ring away? A barrier zone, Wall 4, separates the decayed heartwood from the sound heartwood. Research by investigators at Oxford has shown that barrier zones in red oak contain suberin. It may be that barrier zones separate because they are like an inside bark. This sample shows the highly ordered compartmented make-up of trees. Oak wood is ring porous. It has large spring vessels, and much smaller vessels in the summer. The radial sheets of parenchyma are very pronounced. Note also (at the center) the five-lobed shape of the young oak tree.

Pages 78-79
52. This dissected, experimentally inflicted wound in a sugar maple is very similar to the wound in oak shown in Figure 51. The decayed wood is surrounded by discolored wood. The Walls 2, 3, and 4 are very effective. Maple is a diffuse-porous tree. The spring and summer vessels are the same size, and the vessels are equally distributed in the growth ring. The radial ray sheets are not pronounced as they are in the oak.

53. A natural wound on a large Sitka spruce shows the same pattern as those in the oak and maple. Conifer wood has a different tissue arrangement.

Pages 80-81
54. Tropical hardwoods also follow the CODIT model. The arrow shows the obvious barrier zone. Indeed, microorganisms do not have "free run" in the heartwood. Heartwood does compartmentalize.

55. A pattern similar to that shown in Figure 54 is shown here in an American beech. The central column of discolored wood was sound (A). The column of decayed wood (B) associated with the large branch core wound did not spread into the central column, nor did it spread rapidly into the surrounding columns of discolored wood (C). The CODIT boundaries are not absolute, but they serve as a guide to the spread of microorganisms in trees.
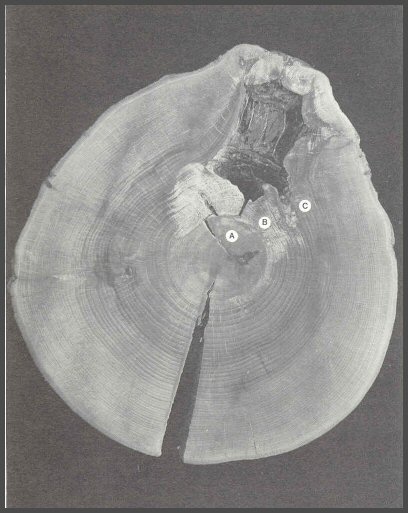
56. The bold barrier zones are easily seen in this mahogany. The barrier zones are often the initial sites for wood separations in living trees and on into wood products.

Pages 82-83
57. The barrier zone may form only a short distance, or it may develop entirely around the tree as shown here in a sweetgum injured by a beaver. The arrow shows the barrier zone. Note also that the insects that infested the decayed wood stayed on the inner side of the barrier zone. The decay spreads as Walls 3 recede. Once the pith is reached by an organism, Wall 2 is no longer in operation. The decay spreads as an opening fan. Why the barrier zone is sometimes extensive and sometimes not, is not well understood. Sometimes even small wounds stimulate large barrier zones, while large wounds may have very small barrier zones. Indeed, much is yet to be learned.

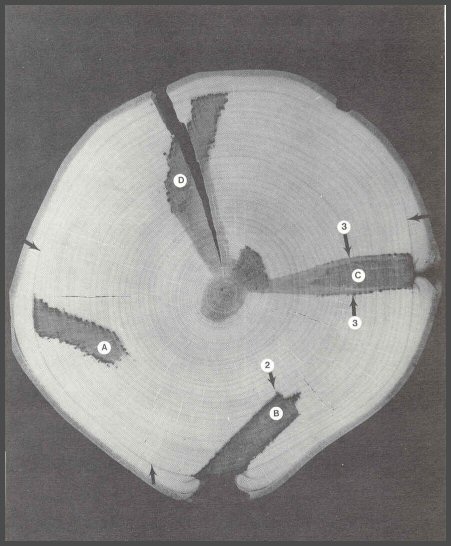
58. The barrier zones associated with the four experimentally inflicted drill bit wounds coalesced to form a complete ring (small arrows). The wood on the inner side of the barrier zone was very slightly darker than the wood that formed after the tree was wounded. Drill wounds A and B were slanted and the internal tips of the wounds were not close to the central column of discolored wood. Drill wounds C and D were close enough to the central column that Walls 2 could not hold: the tissues between the internal tips of the wounds and the central column were isolated, and they died and discolored. Walls 2 did hold for wounds A and B. When new wounds are inflicted in a tree, much that happens internally will depend on what is already in the tree. If the wood is all clear, then the new wounds will be effectively compartmentalized. But when new wounds meet with, or come close to, older inner columns of discolored or decayed wood, compartmentalization may not be as effective. Only Part I of CODIT or Walls 1, 2, and 3, can operate at the time of wounding because the next year's tissues are not there yet. It is possible that Walls 1, 2, and 3 may respond so poorly that the cambium does not survive. Trees do die. Microorganisms do win.
Pages 84-85
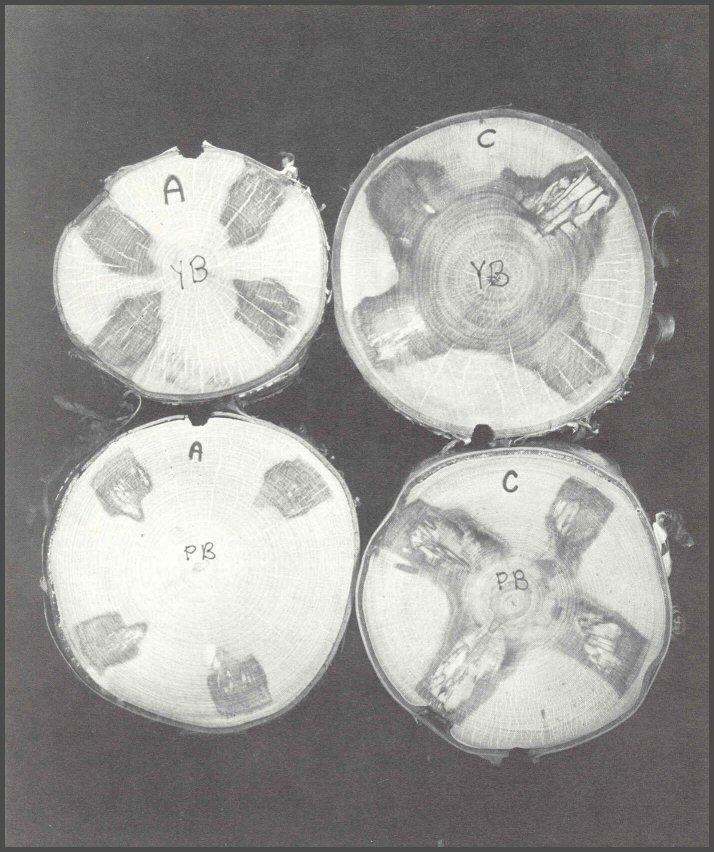
59. Sections from four paper birch (PB) and yellow birch (YB) trees, wounded experimentally with four drill bit wounds each, show the variations in internal response. Tree genetics cannot be discounted. Some individual trees in a species do compartmentalize better than others. Trees YB-A and PB-A were clear to the pith before wounding. This indicates that the trees walled off branch stubs very effectively without having columns of discolored wood spread from the inner branch core wood into the tree trunk. The drill wounds were in clean, healthy tissues, not affected by past columns of discolored wood. The new columns from the drill wounds were effectively compartmentalized. In trees YB-C and PB-C there were large columns of central discolored wood before the drill wounds were inflicted. The large central column suggests that the trees did not effectively wall off discoloration spreading into the trunk from dead branch core wood. The drill holes into trees YB-C and PB-C resulted not only in larger columns of defect, but more decayed wood. Thus the genetics and the wounding history of the tree play major roles in how effectively it compartmentalizes new injuries.
Pages 86-87
60. These samples from the same red maple tree show the single effect of internal defects present when new wounds are made. The red maple received three combination drill-hole and scribe wounds as shown. Wounds A and B had much larger columns of decayed wood than wound C. Why? Wound C was at the base. Studies on many other trees showed that basal wounds usually-but not always-produce smaller columns of decayed wood than wounds higher on the trunk. Bases of most trees are broader than the upper trunks. Also, most columns of central discolored wood associated with dead branch core wood attenuate as they develop downward. So new wounds higher on the trunk usually come closer to older internal columns. Wounds A and B had only a narrow band of healthy wood separating the new wood from the central column. It was not enough to hold Walls 2. Wound C was separated by a large band of clear wood from the very small circle of discolored wood at the pith. Compartmentalization worked very well for wound C.

61. Studies on hybrid poplars were the first to suggest that the degree of compartmentalization may be under genetic control. Fourteen deep drill holes were inflicted in each of many trees, of many different clones. After 2 years the trees were dissected. All trees in some clones had very little discolored wood associated with the wounds, while trees in other clones all had large columns of discolored wood. The two sections shown here are from the best and worst clones. All clones were the same age and they were grown on the same site.

Pages 88-89
62. This peach sample summarizes many of the points made in the group of photos. A discolored column is compartmentalized within the heartwood. A column of decayed and discolored wood is associated with a more recent wound (the hole is where a wood sample was taken for isolation of fungi).

63. Cracks are among the most serious defects in trees. Cracks often start after the trunk has been injured. Oaks commonly have long vertical basal cracks, called "frost cracks". Frost does not start the cracks; wounds do. Old basal fire injuries on oaks are common starting points for cracks.
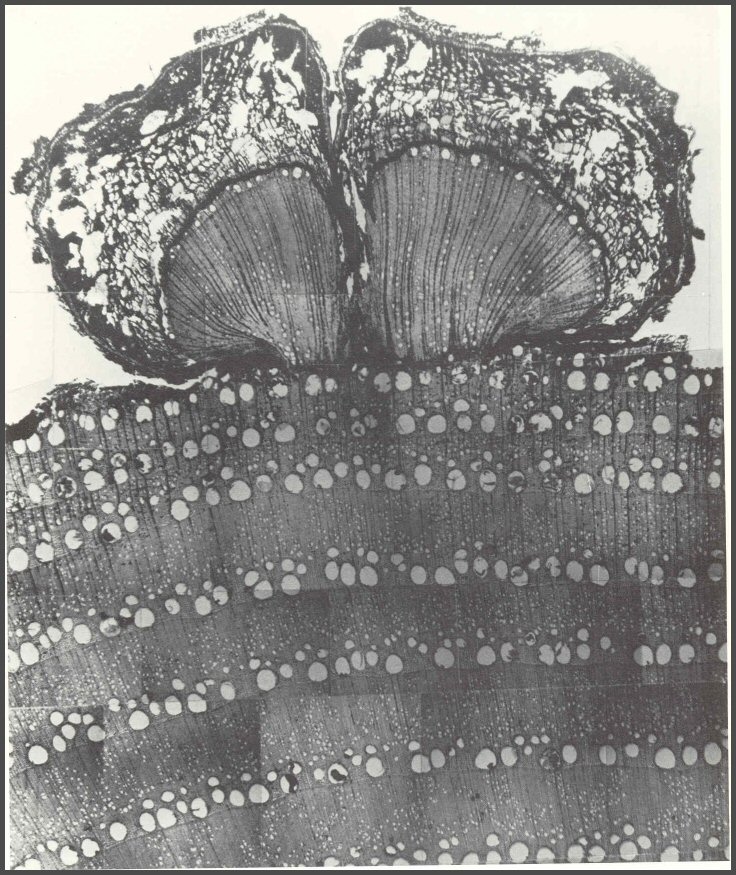
This basal section of red oak shows the internal cracking pattern. The arrows show the circumferential extent of the injury. Note the closure starting with the inroll of the growth rings. Where the inrolling growth rings meet, a primary vertical or radial seam formed. The primary seam is open on this sample. After injury, a Wall 4, or barrier zone, formed, and outward in a radial direction many secondary seams, or shakes formed. Some of the seams split out to the cambium. Often the secondary seams are more serious defects than the primary seam.

Pages 90-91